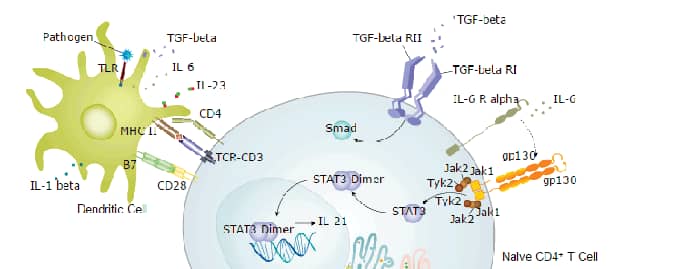
IL-26 was originally cloned from herpesvirus saimiri (HVS)-transformed T-cells and named AK155 (1). It is a member of the IL-10 family of class II cytokines that signal via heterodimeric receptor complexes composed of two type I transmembrane receptor subunits (2). The human IL-26 gene has been mapped to chromosome 12q15. It encodes a 171 amino acid polypeptide with a 21 amino acid signal peptide. In addition to HVS-transformed T cells, IL-26 is also expressed in other virus transformed T cell lines, fresh peripheral mononuclear cells, activated NK cells and T cells. A mouse homologue of human IL-26 has not been identified. IL-26 binds with high-affinity to the heterodimeric complex consisting of the ligand-binding IL-20 R alpha and non ligand-binding IL-10 R beta (3). Activation of the receptor complex results in rapid phosphorylation of STAT1 and STAT3. Although the IL-26 receptor complex is highly specific for IL-26 and is not activated by other class II cytokines, the individual subunits of the IL-26 receptor complex are components in receptor complexes for other class II cytokines (1). IL-20 R alpha can form dimers with IL‑20 R beta to function as signaling receptors for IL-19, IL-20, and IL-24. IL-10 R beta can complex with IL-10 R alpha, IL-22 R, and IL-28 R alpha to transduce signals for IL-10, IL-22, and the three novel IFNs (IL-28A, IL-28B and IL-29), respectively. The physiological functions of IL-26 remain to be determined.
IL-26 was reported to be a homodimer in solution (1). E. coli-derived IL-26 produced by R&D Systems contains disulfide-linked homodimers and monomers. Both the pure monomeric and dimeric IL-26 can bind directly to IL-20 R alpha and induce STAT activation in COLO-205 cells (4). Besides IL-20 R alpha, R&D Systems IL-26 preparations have also been shown to bind IL‑20 R beta. The significance of this receptor-ligand interaction has not been investigated.
 Powered by Bioz
Powered by Bioz