Recombinant Human L1CAM Fc Chimera Protein, CF
R&D Systems | Catalog # 777-NC

Loading...
Key Product Details
- R&D Systems NS0-derived Recombinant Human L1CAM Fc Chimera Protein (777-NC)
- Quality control testing to verify active proteins with lot specific assays by in-house scientists
- All R&D Systems proteins are covered with a 100% guarantee
Source
NS0
Accession Number
Structure / Form
Disulfide-linked homodimer
Applications
Bioactivity
Loading...
Product Specifications
Source
Mouse myeloma cell line, NS0-derived human L1CAM protein
| Human L1CAM (Ile20-Glu1120) & (Arg864âGlu1120) Accession # CAA42508 |
IEGRMD | Human IgG1 (Pro100-Lys330) |
6-His tag |
| N-terminus | C-terminus | ||
Purity
>90%, by SDS-PAGE under reducing conditions and visualized by silver stain.
Endotoxin Level
<0.10 EU per 1 μg of the protein by the LAL method.
N-terminal Sequence Analysis
Ile20 & Arg864
Predicted Molecular Mass
150.4 kDa (monomer) & 55.9 kDa (truncated)
SDS-PAGE
210-225 kDa, reducing conditions
Activity
Measured by the ability of the immobilized protein to support the adhesion of Neuro‑2A mouse neuroblastoma cells.
When 5 x 104 cells/well are added to Recombinant Human L1CAM Fc Chimera coated plates (25 µg/mL with 100 µL/well), >30% will adhere for 1 hour incubation at 37 °C.
Optimal dilutions should be determined by each laboratory for each application.
When 5 x 104 cells/well are added to Recombinant Human L1CAM Fc Chimera coated plates (25 µg/mL with 100 µL/well), >30% will adhere for 1 hour incubation at 37 °C.
Optimal dilutions should be determined by each laboratory for each application.
Reviewed Applications
Read 1 review rated 5 using 777-NC in the following applications:
Formulation, Preparation, and Storage
777-NC
| Formulation | Lyophilized from a 0.2 μm filtered solution in Tris-Citrate and NaCl. |
| Reconstitution | Reconstitute at 200 μg/mL in sterile PBS.
Loading...
|
| Shipping | The product is shipped at ambient temperature. Upon receipt, store it immediately at the temperature recommended below. |
| Stability & Storage | Use a manual defrost freezer and avoid repeated freeze-thaw cycles.
|
Calculators
Background: L1CAM
References
- Maness, P.F. and M. Schachner (2007) Nat. Neurosci. 10:19.
- Wei, C.H. and S.E. Ryu (2012) Exp. Mol. Med. 44:413.
- Faspel, J. and M. Grumet (2003) Front. Biosci. 8:1210.
- Herron, L.R. et al. (2009) Biochem. J. 419:519.
- Rathjen, F.G. and M. Schachner (1984) EMBO. J. 3:1.
- Keifel, H. et al. (2011) Trends Mol. Med. 17:178.
- Dihne, M. et al. (2003) J. Neurosci. 23:6638.
- Kadmon, G. et al. (1998) Dev. Immunol. 6:205.
- Itoh, K. et al. (2000) J. Neurosci. Res. 60:579.
- Jouet, M. et al. (1995) Mol. Brain Res. 30:378.
- He, Q. et al. (2012) Neurosci. Lett. 521:57.
- Thor, G. et al. (1987) EMBO J. 6:2581.
- Sadoul, K. et al. (1988) J. Neurochem. 50:510.
- Hubbe, M. et al. (1993) Eur. J. Immunol. 23:2927.
- Hoja-Lukowicz, D. et al. (2012) Glycoconj J. Apr 29. [Epub ahead of print].
- Geismann, C. et al. (2009) Cancer Res. 69:4517.
- Tischler, V. et al. (2011) Mol. Cancer 10:127.
- Mujoo, K. et al. (1986) J. Biol. Chem. 261:10299.
- Kobayashi, M. et al. (1991) Biochim. Biophys. Acta 1090:238.
- Hlavin, M.L. and V. Lemmon (1991) Genomics 11:416.
- Castellani, V. et al. (2002) EMBO J. 21:6348.
- Zhao, X. and C-H Siu (1995) J. Biol. Chem. 270:29413.
- Oleszewski, M. et al. (1999) J. Biol. Chem. 274:24602.
- Felding-Habermann, B. et al. (1997) J. Cell Biol. 139:1567.
- Kamiguchi, H. et al. (1998) J. Neurosci. 18:5311.
- De Angelis, E. et al. (2001) J. Biol. Chem. 276:32738.
- Montgomery, A.M.P. et al. (1996) J. Cell Biol. 132:475.
- Sadoul, R. et al. (1989) J. Neurochem. 53:1471.
- Reidle, S. et al. (2009) Biochem. J. 420:391.
- Lutz, D. et al. (2012) J. Biol. Chem. 287:17161.
Long Name
Cell Adhesion Molecule L1
Alternate Names
CAML1, CD171, HSAS, HSAS1, MASA, MIC5, NCAM-L1, S10, SPG1
Gene Symbol
L1CAM
UniProt
Additional L1CAM Products
Product Documents for Recombinant Human L1CAM Fc Chimera Protein, CF
Certificate of Analysis
To download a Certificate of Analysis, please enter a lot or batch number in the search box below.
Note: Certificate of Analysis not available for kit components.
Product Specific Notices for Recombinant Human L1CAM Fc Chimera Protein, CF
For research use only
Citations for Recombinant Human L1CAM Fc Chimera Protein, CF
 Powered by Bioz
Powered by Bioz
Customer Reviews for Recombinant Human L1CAM Fc Chimera Protein, CF (1)
5 out of 5
1 Customer Rating
Have you used Recombinant Human L1CAM Fc Chimera Protein, CF?
Submit a review and receive an Amazon gift card!
$25/€18/£15/$25CAN/¥2500 Yen for a review with an image
$10/€7/£6/$10CAN/¥1110 Yen for a review without an image
Submit a review
Customer Images
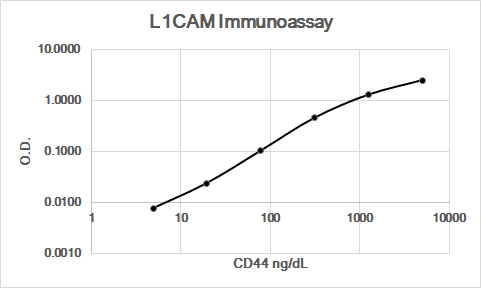
Showing
1
-
1 of
1 review
Showing All
Filter By:
-
Application: Immunoassay StandardVerified Customer | Posted 06/17/2019

There are no reviews that match your criteria.
Loading...