Differentiation Potential of Induced Pluripotent Stem Cells
Somatic cells that have been reprogrammed to a pluripotent state, known as induced pluripotent stem (iPS) cells, are generating excitement due to their ability to function like embryonic stem (ES) cells.1, 2 Unlike ES cells, iPS cells are more readily obtainable for therapy and research, and their isolation does not carry the same ethical concerns.1, 2 Human iPS cells may be an ideal source for patient-specific therapy since they can be derived from the patients themselves. In addition, iPS cells can serve as useful research tools by providing: 1) models of human disease to use for screening new drugs or for studying mechanisms of pathogenesis and toxicology, and 2) models of normal development to use for screening potential teratogens or for understanding tissue repair and regeneration.1, 3
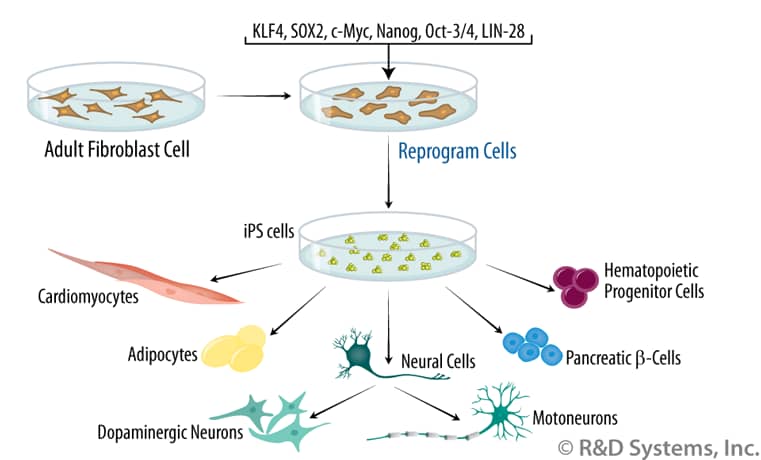
Directed Differentiation of iPS Cells. Induced pluripotent stem cells (iPS cells) are generated by reprogramming adult somatic cells. Following isolation, somatic cells are cultured in vitro and transduced with expression vectors encoding transcription factors associated with pluripotency. For most cell types, four factors (c-Myc, Oct-3/4, SOX2, and KLF4) are used, although a combination of alternate factors (Oct-3/4, SOX2, Nanog, and LIN-28) has also been used successfully. Expression of these exogenous factors triggers a gradual process of silencing markers of the differentiated phenotype and inducing markers of the pluripotent state in some cells. As pluripotent cells, iPS cells theoretically have the ability to generate all cell types found in the body. Human iPS cells have been differentiated into a variety of these, including adipocytes, cardiomyocytes, primitive hematopoietic cells, pancreatic beta-cells, and several different neuronal cell types. [Figure adapted from Amabile, G. & A. Meissner (2009) Trends Mol. Med. 15:59.]
iPS cells were first generated by Takahashi and Yamanaka using mouse fibroblasts that had been transduced with four transcription factors: Oct-3/4, SOX2, c-Myc, and KLF4, under ES cell culture conditions.2 These cells exhibit the morphology, growth, marker expression, and pluripotency of ES cells.2, 3 Further research demonstrated the ability to generate human pluripotent stem cells from adult fibroblasts. Interestingly, there is some flexibility in which transcription factors can be used to reprogram cells. While one report used the same four transcription factors as Takahashi and Yamanaka had used in mice, another study used Oct-4, SOX2, Nanog, and LIN-28 for reprogramming human somatic cells.4, 5 iPS cells appear to exhibit many of the same outward characteristics as ES cells including morphology, proliferation, epigenetic status, and pluripotency.4, 5 However, expression array analysis does suggest that differences in gene expression signatures exist between the cell types.6 How these differences might affect iPS cells phenotypically is a matter of ongoing research.
Induced Pluripotent Stem Cells from Mice
Recently, researchers have successfully created mice from iPS cells.7, 8 The success rate for implantation of embryos resulting in viable mice was in the low single digits, and a number of iPS-derived mice exhibited physical abnormalities. Nonetheless, some survived into adulthood and produced viable offspring. In human cells, researchers have demonstrated the capability and efficacy of iPS cells to differentiate into a variety of cell types. Using four different human iPS cell lines, Taura et al. were able to show an adipogenic potential comparable to human ES cells.9 These iPS cells exhibited lipid accumulation and expression of adipogenesis markers including C/EBP alpha, PPAR gamma, Leptin, and FABP4. Research from another group showed that iPS cells could be used to generate CD34+CD43+ hematopoietic progenitors and CD31+CD43- endothelial cells under conditions similar to that used for ES cells.10 These human iPS-derived cells could be further separated into phenotypically defined subsets of primitive hematopoietic cells in a pattern of differentiation resembling that of ES cells. Human iPS cell lines were also induced to differentiate into pancreatic insulin-producing cells.11 After first generating PDX-1 positive progenitor cells, human iPS cells were further differentiated into pancreatic cells expressing MafA, Glut2, insulin, and in some cases, amylase and C-peptide. Functional cardiomyocytes demonstrating sarcomeric organization and expressing cardiac markers including Nkx2.5, cardiac Troponin T, atrial natriuretic factor, and myosin heavy and light chains, were also derived from human iPS cells and were indistinguishable from those generated from ES cells.12 Electrophysiology experiments revealed that like ES cells, iPS cells differentiate into nodal-, atrial-, and ventricular-like phenotypes, and are responsive to the canonical cardiomyocyte beta-adrenergic signaling pathway. Both human ES and iPS cells were efficiently converted to neural cells using two inhibitors of TGF-beta/Smad signaling, Noggin and the drug SB431542.13 The synergistic action of these two inhibitors resulted in Pax6+ primitive neural cells that could then be further differentiated into the neural crest, anterior CNS, somatic motoneurons, and dopaminergic neurons.
These examples of iPS cells differentiating into various somatic cell types pave the way for generating patient-specific pluripotent cells that could be used for research and therapy. Disease research, drug development, and patient therapy will be greatly enhanced by the ability to recapitulate normal and pathologic tissue differentiation and formation in vitro, which iPS cells provide more readily than ES cells.
References
- Nishikawa, S. et al. (2008) Nat. Rev. Mol. Cell Biol. 9:725.
- Takahashi, K. & S. Yamanaka (2006) Cell 126:663.
- Amabile, G. & A. Meissner (2009) Trends Mol. Med. 15:59.
- Takahashi, K. et al. (2007) Cell 131:861.
- Yu, J. et al. (2007) Science 318:1917.
- Chin, M.H. et al. (2009) Cell Stem Cell 5:111.
- Zhao, X-Y. et al. (2009) Nature. Advanced online publication.
- Kang, L. et al. (2009) Cell Stem Cell. 5:135.
- Taura, D. et al. (2009) FEBS Lett. 583:1029.
- Choi, K-D. et al. (2009) Stem Cells 27:559.
- Zhang, D. et al. (2009) Cell Res. 19:429.
- Zhang, J. et al. (2009) Circ. Res. 104:e30.
- Chambers, S.M. et al. (2009) Nat. Biotechnol. 27:275.
This symbol denotes references that cite the use of R&D Systems products.
Related Information
Human Pluripotent Stem Cell Antibody Array
Interactive Pathway: Embryonic and Induced Pluripotent Stem Cells & Lineage-specific Markers
New ExCellerate™ iPSC Expansion Medium
Technical Notes: Assessing the Pluripotent Status of Stem Cells