(+)-MK 801 maleate
Tocris Bioscience | Catalog # 0924
-MK 801 maleate | CAS No. 77086-22-7")
Key Product Details
Description
Alternative Names
Product Description
(+)-MK 801 maleate is a high affinity (Ki = 37.2 nM), selective and non-competitive NMDA receptor antagonist. (+)-MK 801 maleate acts by binding to a site located within the NMDA associated ion channel and thus prevents Ca2+ flux. It is an effective anti-ischemic agent in several animal models. (+)-MK 801 maleate increases motor activity in rat models. (+)-MK 801 maleate inhibits proliferation and increases apoptosis in hippocampal neural stem cells (NSCs). (+)-MK 801 maleate induces schizophrenia-like symptoms in rodents.(-)-enantiomer also available.
Product Specifications for (+)-MK 801 maleate
Molecular Weight
Formula
Storage
Purity
Chemical Name
CAS Number
PubChem ID
InChI Key
SMILES
The technical data provided above is for guidance only. For batch specific data refer to the Certificate of Analysis.
Solubility
| Solvent | Max Conc. mg/mL | Max Conc. mM | |
|---|---|---|---|
| Solubility | |||
| water | 3.37 | 10 with gentle warming | |
| DMSO | 33.74 | 100 |
Preparing Stock Solutions for (+)-MK 801 maleate
The following data is based on the product molecular weight 337.37.
Batch specific molecular weights may vary from batch to batch due to the degree of hydration, which all affect the solvent volumes required to prepare stock solutions.
| Concentration / Solvent Volume / Mass | 1 mg | 5 mg | 10 mg |
|---|---|---|---|
| 1 mM | 2.96 mL | 14.82 mL | 29.64 mL |
| 5 mM | 0.59 mL | 2.96 mL | 5.93 mL |
| 10 mM | 0.30 mL | 1.48 mL | 2.96 mL |
| 50 mM | 0.06 mL | 0.30 mL | 0.59 mL |
Calculators
Background References
References are publications that support the biological activity of the product. See our Citations tab to view 120 publications citing the usage of this product.
- Ding Effect of NMDA on proliferation and apoptosis in hippocampal neural stem cells treated with MK-801. Exp.Ther.Med. 2018 PMID: 30116364
- Carey Effects of dizocilpine (MK-801) on motor activity and memory. Psychopharmacology 1998 PMID: 9683001
- Zajaczkowski Uncompetitive NMDA receptor antagonists attenuate NMDA-induced impairment of passive avoidance learning and LTP. Neuropharmacology 1997 PMID: 9257940
- Wong The anticonvulsant MK 801 is a potent N-MthD.-aspartate antagonist. Proc.Natl.Acad.Sci.U.S.A. 1986 PMID: 3529096
- Hatfield The dose-response relationship and therapeutic window for dizocilpine (MK-801) in a rat focal ischaemia model. Eur.J.Pharmacol. 1992 PMID: 1526248
- Gill The neuroprotective action of dizocilpine (MK-801) in the rat middle cerebral artery occlusion model of focal ischaemia. Br.J.Pharmacol. 1991 PMID: 1912992
Product Documents for (+)-MK 801 maleate
Certificate of Analysis
To download a Certificate of Analysis, please enter a lot or batch number in the search box below.
Product Specific Notices for (+)-MK 801 maleate
For research use only
Related Research Areas
Citations for (+)-MK 801 maleate
 Powered by Bioz
Powered by Bioz
Customer Reviews for (+)-MK 801 maleate (1)
Have you used (+)-MK 801 maleate?
Submit a review and receive an Amazon gift card!
$25/€18/£15/$25CAN/¥2500 Yen for a review with an image
$10/€7/£6/$10CAN/¥1110 Yen for a review without an image
Submit a review
Customer Images
-
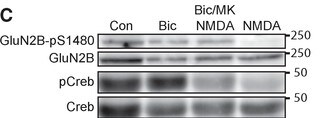
Species: RatAssay Type: In VitroCell Line/Tissue: Sprague-Dawley rats primary neuronalVerified Customer | Posted 04/04/202010 μM MK-801 for 10 min

There are no reviews that match your criteria.