MNI-caged-L-glutamate
Tocris Bioscience | Catalog # 1490

Key Product Details
Description
Alternative Names
Product Description
MNI-caged-L-glutamate is a form of glutamate linked to a photo-protecting group, 4-methoxy-7-nitroindolinyl (MNI); it rapidly and efficiently releases L-glutamate (Cat. No. 0218) by photolysis (300 - 380 nm excitation) with a quantum yield in the 0.065-0.085 range. It is also suitable for use with two-photon uncaging microscopy (cross-section of 0.06 GM at 730 nm). MNI-caged-L-glutamate is optically compatible with other chromophores used for fluorescence imaging, such as GFP, YFP and most Ca2+ dyes. MNI-caged-L-glutamate is 2.5-fold more efficient at releasing L-glutamate than NI-caged L-glutamate. MNI-caged-L-glutamate is water-soluble, stable at neutral pH, highly resistant to hydrolysis and pharmacologically inactive at neuronal glutamate receptors and transporters (up to mM concentrations). MNI-caged-L-glutamate can be used for in situ studies of fast synaptic glutamate receptors.View more information regarding MNI-caged-L-glutamate.
Licensing Information
Sold under license from the Medical Research Council
Product Specifications for MNI-caged-L-glutamate
Molecular Weight
Formula
Storage
Purity
Chemical Name
CAS Number
PubChem ID
InChI Key
SMILES
The technical data provided above is for guidance only. For batch specific data refer to the Certificate of Analysis.
Solubility
| Solvent | Max Conc. mg/mL | Max Conc. mM | |
|---|---|---|---|
| Solubility | |||
| water | 16.16 | 50 |
Preparing Stock Solutions for MNI-caged-L-glutamate
The following data is based on the product molecular weight 323.30.
Batch specific molecular weights may vary from batch to batch due to the degree of hydration, which all affect the solvent volumes required to prepare stock solutions.
| Concentration / Solvent Volume / Mass | 1 mg | 5 mg | 10 mg |
|---|---|---|---|
| 0.5 mM | 6.19 mL | 30.93 mL | 61.86 mL |
| 2.5 mM | 1.24 mL | 6.19 mL | 12.37 mL |
| 5 mM | 0.62 mL | 3.09 mL | 6.19 mL |
| 25 mM | 0.12 mL | 0.62 mL | 1.24 mL |
Calculators
Background References
References are publications that support the biological activity of the product. See our Citations tab to view 221 publications citing the usage of this product.
- Ellis-Davies Two-Photon Uncaging of Glutamate Front Synaptic Neurosci. 2019 PMID: 30687075
- Palma-Cerda New caged neurotransmitter analogs selective for glutamate receptor sub-types based on methoxynitroindoline and nitrophenylethoxycarbonyl caging groups. Neuropharmacology. 2012 PMID: 22609535
- Papageorgiou and Corrie Effects of aromatic substitutions on the photocleavage of 1-acyl-7-nitroindolines. Tetrahedron
- Matsuzaki Dendritic spine geometry is critical for AMPA receptor expression in hippocampal CA1 pyramidal neurons. Nat.Neurosci. 2001 PMID: 11687814
- Maier Comparative analysis of inhibitory effects of caged ligands for the NMDA receptor. J.Neurosci.Methods 2005 PMID: 15652611
- Canepari Photochemical and pharmacological evaluation of 7-nitroindolinyl- and 4-methoxy-7-nitroindolinyl-amino acids as novel, fast caged neurotransmitters. J.Neurosci.Methods 2001 PMID: 11640955
Product Documents for MNI-caged-L-glutamate
Certificate of Analysis
To download a Certificate of Analysis, please enter a lot or batch number in the search box below.
Product Specific Notices for MNI-caged-L-glutamate
For research use only
Citations for MNI-caged-L-glutamate
 Powered by Bioz
Powered by Bioz
Customer Reviews for MNI-caged-L-glutamate (1)
Have you used MNI-caged-L-glutamate?
Submit a review and receive an Amazon gift card!
$25/€18/£15/$25CAN/¥2500 Yen for a review with an image
$10/€7/£6/$10CAN/¥1110 Yen for a review without an image
Submit a review
Customer Images
-
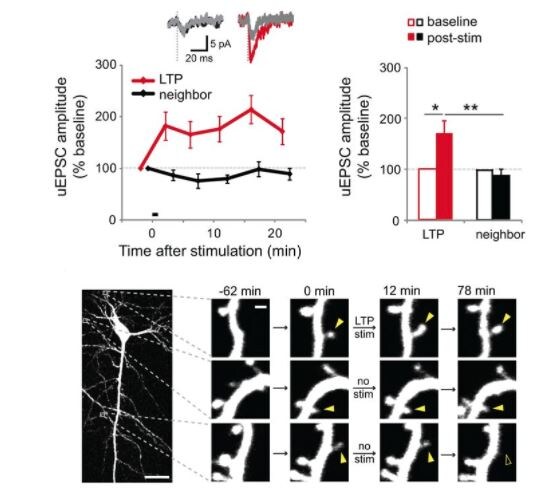
Assay Type: In VitroCell Line/Tissue: Organotypic Hippocampus SlicesVerified Customer | Posted 10/20/2017We resuspend in ACSF and found that we can freeze and reuse the solution. However, when suspended in HBSS it is no longer effective after freezing.We buy this product in bulk for use in all of our uncaging experiments. High frequency uncaging next to dendritic spine head induces reliable spine growth, indicative of synapse strengthening.

There are no reviews that match your criteria.