Cytokines and Receptors Research

Cytokine Signaling Regulates Multiple Biological Processes
Cytokines are small extracellular signaling molecules secreted by cells that function as intercellular messengers to regulate a wide range of biological processes. They exert their effects by binding to specific cell surface cytokine receptors, which are often multi-subunit transmembrane proteins. Upon ligand binding, these receptors typically dimerize or oligomerize, initiating intracellular signal transduction cascades such as the Janus kinase (Jak)-STAT, MAPK/ERK, PI3-K/Akt, and IκB/NF-κB pathways. Activation of these pathways triggers phosphorylation events that stimulate transcription factors, which in turn regulate gene expression programs controlling fundamental cellular processes including:
- Cell activation
- Proliferation
- Differentiation
- Migration
- Survival
- Apoptosis
Cytokines, including colony-stimulating factors (e.g., G-CSF, GM-CSF), interferons (IFN-gamma), and tumor necrosis factor alpha (TNF-alpha), are produced by various immune (macrophages, dendritic cells, T cells, B cells, monocytes, neutrophils, eosinophils) and non-immune cells (fibroblasts, epithelial cells, stromal cells, endothelial cells, stem cells). Production varies by cell type; For example, IFN-gamma is mainly secreted by activated T cells and natural killer cells, while G-CSF and GM-CSF are primarily produced by macrophages and stromal cells. Collectively, cytokines regulate processes such as hematopoiesis and orchestrate inflammatory responses essential for immune system function and wound healing. Paradoxically, they can also drive the development of chronic inflammatory and autoimmune diseases, including rheumatoid arthritis, multiple sclerosis, inflammatory bowel disease, psoriasis, and type I diabetes.
R&D Systems Rigorous In-House Testing Ensures the Performance and Consistency of Our Cytokine Research Reagents
R&D Systems has a rich history in the field of cytokine biology. We purified and sold our first recombinant cytokines in 1988. Since that time, we have continued to build our cytokine catalog and have used these rigorously tested proteins to produce high quality antibodies and the most trusted and widely published cytokine ELISA Kits on market.
Our ability to develop and manufacture over 95% of our proteins, antibodies, and immunoassays in-house is a core capability that sets us apart from the competition. It gives us remarkable control over reagent optimization during product development, and it allows us to implement quality tests that are unavailable to most reagent distributors. This ensures that we are providing our customers with the highest performing and most consistent reagents in the biomedical research market.
R&D Systems Product Development Workflow
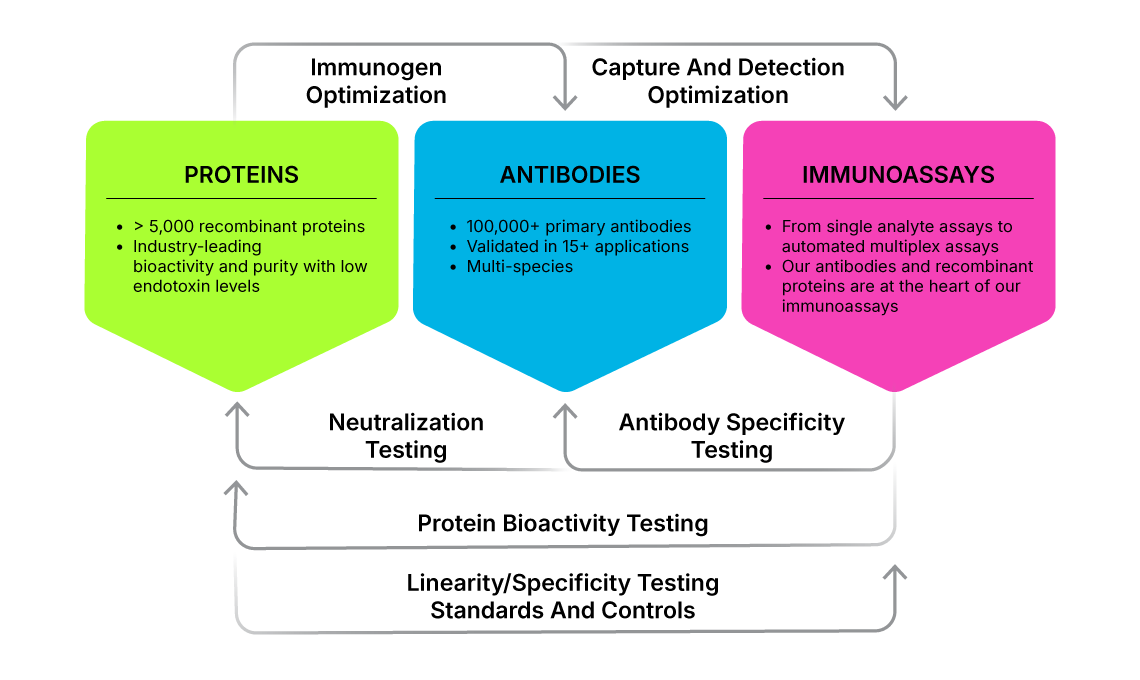
R&D Systems Product Development Workflow and Rigorous In-House Testing Produces High Quality Cytokines, Antibodies, and Immunoassays. More than 95% of the 5,000 proteins offered by R&D Systems are manufactured in-house and undergo our rigorous quality testing to ensure high activity, purity, and lot-to-lot consistency. Using these high quality proteins as immunogens, we generate antibodies, making us one of the few antibody suppliers that still develops antibodies in-house. This in-house development, coupled with our strict application-focused antibody validation process and quality control specifications ensure exceptional antibody specificity and performance. R&D Systems' antibodies, along with our own proteins and diluents are then used to build R&D Systems ELISA Kits, the most trusted and widely published ELISA Kits on market. Our ELISA Kits are developed by testing several different monoclonal and polyclonal antibodies to determine which combination optimally couples for use in analyte detection. Selected antibodies are then carefully titered to ensure that the concentrations chosen will give the best possible results for the assay. Finally, the complete ELISA Kit is carefully tested to confirm that it meets our rigorous standards for specificity, recovery, linearity, and precision.
Cytokines for Cell Culture
Cytokines are some of the most important components of cell cultures as they are necessary to maintain healthy cells, drive cell proliferation, and promote differentiation into specific cell lineages. R&D Systems rigorous in-house testing and quality control specifications ensure that our proteins will provide the highest levels of activity, purity, and lot-to-lot consistency, so that you can quickly standardize your protocols and minimize variability from one experiment to the next.
- Stringent quality control specifications for bioactivity, purity, and endotoxin levels
- Lot-to-lot consistency ensured by testing each new lot side-by-side with previous lots
- Large selection that includes over 5,000 bioactive, research-grade, Animal-free Preclinical, and GMP-grade proteins to address researchers’ needs from basic research to clinical manufacturing
Antibodies are essential tools for detecting and analyzing the functions of different cytokines. Our catalog includes thousands of antibodies validated for specificity and reproducibility in one or more of 15 applications, including blocking/neutralization, flow cytometry, immunocytochemistry, immunohistochemistry, Western blot, and Simple Western™.
- Extensive in-house validation and expert product support ensure performance
- Data images, citations, reviews, and a 100% satisfaction guarantee provide confidence that our antibodies will work in the applications and species designated
- Over 1,400 blocking/neutralizing antibodies validated with the same bioassays used to test the bioactivity of our proteins, ensuring they block/neutralize the appropriate cellular functions
- A wide assortment of primary antibodies, coupled with a selection of over 30 fluorescent labels, makes it easier than ever to find the exact antibody-fluorochrome combinations you need to directly detect your targets of interest, and provides the highest levels of flexibility for multiplex experiments

For cytokine profiling and analysis, we offer a wide variety of immunoassays for either single- or multianalyte quantification.
- Single analyte ELISA kit formats include our ready-to-use Quantikine™ ELISAs, rapid 90-minute QuicKit™ ELISAs, and the flexible DuoSet™ ELISA Development Systems that provide the key components necessary for a customer to develop their own customized assay.
- Multiplex options include Simple Plex™ Assays on the Ella™ System and bead-based Luminex® Assays.
- Simple Plex assays on the Ella System offer rapid, hands-free multiplex testing for up to 8 analytes. This fully automated, microfluidic cartridge-based system will provide reproducible results in under 90 minutes.
- For enhanced multiplexing capabilities, Luminex assays provide one of the largest mix-and-match menus available, enabling precise and accurate quantification of up to 50 analytes simultaneously.
- Chemokines
- Common beta chain receptor family
- Common gamma chain receptor family
- Interleukin-1 (IL-1) family cytokines
- Interleukin-6 (IL-6) family cytokines
- Interleukin-10 (IL-10) family cytokines
- Interleukin-12 (IL-12) family cytokines
- Interleukin-17 (IL-17) family cytokines
- Interferons
- TNF superfamily cytokines
Given the critical role of cytokine-receptor interactions in immune regulation, strategies to selectively modulate these signaling events have become a focal point in therapeutic development. Cytokine-cytokine receptor binding specificity enables binding proteins and receptor antagonists to competitively inhibit cytokine signaling, thereby modulating key signal transduction pathways. This modulation is critical for maintaining immune homeostasis and preventing dysregulated inflammatory responses. Therapeutic strategies employing inhibitors and monoclonal antibodies targeting cytokine receptors or downstream signaling components, including tyrosine kinases, have emerged as essential tools for regulating inflammatory cytokine activity across diverse disease contexts.
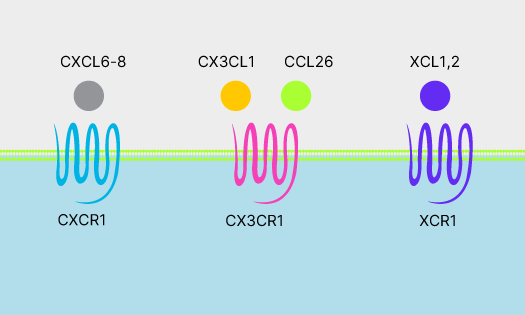
Chemokines
Chemokines regulate immune cell migration under homeostatic and inflammatory conditions. Learn about chemokine subfamilies and functions.
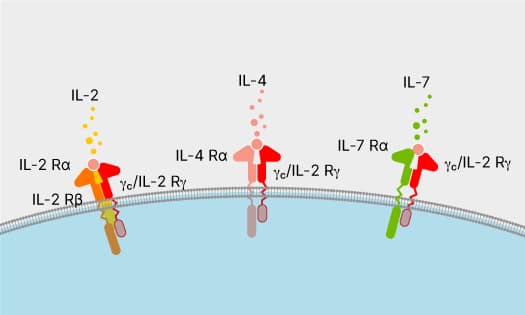
Common Receptor Gamma Chain Family
Common receptor gamma chain cytokines regulate the development and functions of immune cells. Read more about common gamma chain family cytokines.
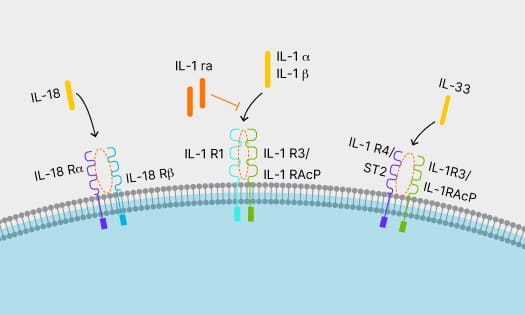
IL-1 Family Cytokines
IL-1 family cytokines have pro- and anti- inflammatory effects. Learn more about IL-1 family cytokines and signaling pathways.

IL-6 Family Cytokines
IL-6 family cytokines have both unique and overlapping functions on hematopoietic cells. Explore the functions of IL-6 family cytokines.

IL-10 Family Cytokines
Despite similarities in their receptor complexes and signaling pathways, IL-10 family cytokines have diverse biological effects. Learn more.

IL-12 Family Cytokines
The IL-12 family consists of structurally similar heterodimeric cytokines with distinct immune regulatory effects. Read about the IL-12 Family.

IL-17 Family Cytokines
IL-17 family cytokines are produced by multiple cell types and primarily have pro-inflammatory effects. Explore IL-17 signaling pathways.

Interferons
Interferons have anti-viral and immune regulatory effects. Discover the signaling pathways activated by type I,
type II, and type III IFNs.

TNF Superfamily
TNF superfamily proteins regulate the functions of innate and adaptive immune cells. Learn more about the TNF superfamily.
Additional Cytokine Products for Cell Culture and Analysis
Animal-free Recombinant Proteins
Animal-free proteins provide a seamless transition from preclinical research into clinical manufacturing, ensure defined culture conditions, and eliminate regulatory or ethical concerns associated with animal-derived reagents. Our Animal-freee proteins are produced using identical expression systems and manufacturing methods as our Animal-free GMP-grade proteins.
GMP-grade Proteins
GMP-grade proteins are manufactured under guidelines that allow for their use as ancillary materials in cell therapy manufacturing processes. These proteins undergo extensive quality control testing and come with comprehensive documentation and full transparency and traceability of source and manufacturing system. Liquid, process-sized, and closed process options are also available for GMP cytokines. Learn more.
Cytokine Signaling Inhibitors
Tocris small molecule cytokine receptor and signaling pathway inhibitors enable detailed investigation of the pathways that control immune activation, inflammation, and disease. Targeting key cascades like Jak-STAT, IkB/NFkB, and MEK/ERK, these modulators support research on cell proliferation, differentiation, apoptosis, and immune responses to help advance target validation and our understanding of cytokine biology and disease models.
ELISpot Kits
ELISpot Kits are highly sensitive, microplate-based assays for the detection of cytokine-secreting cells. Complete ELISpot Kits are ready-to-run and require no assay development or refinement. As an alternative to our complete kits, we also offer ELISpot Development Modules, which provide a flexible, do-it-yourself format for ELISpot development.
Proteome Profiler™ Antibody Arrays
Proteome Profiler Antibody Arrays are membrane-based arrays designed to simultaneous measure up to 119 proteins in a single sample. They provide a simple, economical, semi-quantitative multiplex assay for early stage discovery research.
Luminex® Assays
Luminex panels are bead-based multiplex assays that enable you to multiplex up to 50 analytes, conserving on sample, time, and cost. Select from more than 490 targets spanning human, mouse, rat, porcine, and non-human primate species. Luminex Assays will deliver data that is reliable and reproducible because every panel is QC tested.
Custom Services
If you can’t find the protein, antibody, or immunoassay that you need on our website, contact us. From scratch development to customizing a reagent from our catalog, our experienced custom services team will work with you to create a reagent that fits your experimental needs.
Periodic Table of Cytokine Families Poster
We love to make and measure cytokines and combine them in interesting ways. Request our cytokine families poster to brighten up your lab.
Proteins Designed for Performance
AI Modified Proteins are engineered to address cell culture challenges, with tailored functionalities to improve cell therapies.
ELISA Guide to Successful Experiments
Explore our comprehensive guide to running a successful ELISA, including different formats, best practices, controls, and troubleshooting.