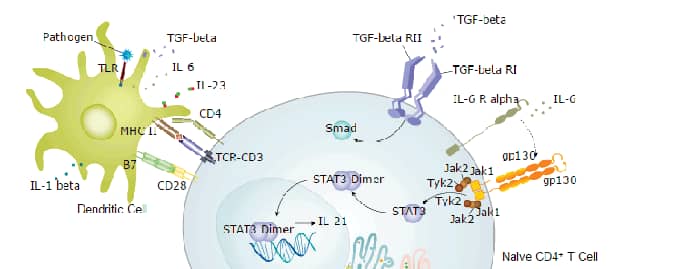
Interleukin 17 (IL-17; also IL-17A and CTLA-8) is a 17 kDa member of the IL-17 family of cytokines (1). Members of this family demonstrate a structural motif termed a cysteine knot which characterize a large superfamily of growth factors. Although most cysteine knot superfamily members use three intrachain disulfide bonds to create a knot, IL-17 family molecules generate the same structural form with only two disulfide links (2-4). Based on the amino acid (aa) sequence alignment with human IL-17, canine IL-17 is 130 aa in length. It is secreted as a 35 kDa disulfide-linked homodimer and as a 40 kDa disulfide-linked heterodimer with IL-17F (5). Canine IL‑17 is 81% identical on the aa level to human IL-17. IL-23 drives Th17 lymphocytes to produce IL-17 (6-8). IL-17’s production has also been demonstrated in gamma delta T cells (9), CD8+ memory T cells (10-11), eosinophils (12), neutrophils (10), and monocytes (13). Studies have identified that the widely expressed receptors IL‑17RA and IL-17RC form a heterodimer for the binding of IL-17 (6, 14-15). The predominant function of IL-17 is thought to be as a proinflammatory mediator through a variety of mechanisms (16). Locally, IL‑17 stimulates production of IL-6, prostaglandin E and nitric oxide (16-19), and synergy with other inflammatory cytokines such as TNF-alpha, IL‑1 beta and IFN -gamma leads to up-regulation of gene expression and progression and amplification of local inflammation (16, 20-22). IL‑17 also mediates chemotaxis of neutrophils and monocytes to sites of inflammation through the chemoattractant mediators IL-8, GRO‑ alpha, and MCP-1 (16, 22-25) while augmenting production of hematopoietic growth factors, such as G-CSF and GM-CSF (16, 26, 27), which promote the growth and maturation of the recruited myeloid cells. In addition, IL-17 serves as a bridge between innate and adaptive immune responses by enhancing the induction of co-stimulatory molecules such as ICAM-1 and other cytokines (16, 22, 28), thereby supporting T cell activation. IL-17 expression has been associated with many inflammatory diseases, such as rheumatoid arthritis, multiple sclerosis, asthma, systemic lupus erythematosus and allograft rejection (15).
Canine IL-17/IL-17A Antibody (665909)
R&D Systems | Catalog # MAB5848

Key Product Details
Species Reactivity
Canine
Applications
Neutralization
Label
Unconjugated
Antibody Source
Monoclonal Mouse IgG2A Clone # 665909
Loading...
Product Specifications
Immunogen
E. coli-derived recombinant canine IL-17
Gly26-Ala155
Accession # NP_001159350
Gly26-Ala155
Accession # NP_001159350
Specificity
Detects canine IL-17 in direct ELISAs. In direct ELISAs, approximately 25%
cross-reactivity with recombinant human (rh) IL-17A is observed and no
cross-reactivity with rhIL-17F or recombinant mouse IL-17A is observed.
Clonality
Monoclonal
Host
Mouse
Isotype
IgG2A
Endotoxin Level
<0.10 EU per 1 μg of the antibody by the LAL method.
Scientific Data Images for Canine IL-17/IL-17A Antibody (665909)
IL-6 Secretion Induced by IL‑17 and Neutralization by Canine IL‑17 Antibody.
Recombinant Canine IL-17 (Catalog # 5848-CL) induces IL-6 secretion in the NIH-3T3 mouse embryonic fibroblast cell line in a dose-dependent manner (orange line). IL-6 Secretion elicited by Recombinant Canine IL-17 (5 ng/mL) is neutralized (green line) by increasing concentrations of Mouse Anti-Canine IL-17 Monoclonal Antibody (Catalog # MAB5848). The ND50 is typically 5-30 ng/mL.Applications for Canine IL-17/IL-17A Antibody (665909)
Application
Recommended Usage
Neutralization
Measured by its ability to neutralize IL‑17-induced IL-6 secretion in the NIH‑3T3 mouse embryonic fibroblast cell line. Yao, Z. et al. (1995) Immunity 3:811.The Neutralization Dose (ND50) is typically 5-30 ng/mL in the presence of 5 ng/mL Recombinant Canine IL‑17.
Formulation, Preparation, and Storage
Purification
Protein A or G purified from hybridoma culture supernatant
Reconstitution
Sterile PBS to a final concentration of 0.5 mg/mL. For liquid material, refer to CoA for concentration.
Loading...
Formulation
Lyophilized from a 0.2 μm filtered solution in PBS with Trehalose. *Small pack size (SP) is supplied either lyophilized or as a 0.2 µm filtered solution in PBS.
Shipping
Lyophilized product is shipped at ambient temperature. Liquid small pack size (-SP) is shipped with polar packs. Upon receipt, store immediately at the temperature recommended below.
Stability & Storage
Use a manual defrost freezer and avoid repeated freeze-thaw cycles.
- 12 months from date of receipt, -20 to -70 °C as supplied.
- 1 month, 2 to 8 °C under sterile conditions after reconstitution.
- 6 months, -20 to -70 °C under sterile conditions after reconstitution.
Calculators
Background: IL-17/IL-17A
References
- Gaffen, S.L. et al. (2006) Vitam. Horm. 74:255.
- Kawaguchi, M. et al. (2004) J. Allergy Clin. Immunol. 114:1265.
- Kolls, J.K. and A. Linden (2004) Immunity 21:467.
- Moseley, T.A. et al. (2003) Cytokine Growth Factor Rev. 14:155.
- Wright, J.F. et al. (2007) J. Biol. Chem. 282:13447.
- Cheung, P.F.Y. et al. (2008) J. Immunol. 180:5625.
- Steinman, L. (2007) Nat. Med. 13:139.
- Hunter, C.A. (2005) Nat. Rev. Immunol. 5:521.
- Lockhart, E. et al. (2006) J. Immunol. 177:4662.
- Ferretti, S. et al. (2003) J. Immunol. 170:2106.
- Shin, H.C. et al. (1999) Cytokine 11:257.
- Molet, S. et al. (2001) J. Allergy Clin. Immunol. 108:430.
- Zhou, Q. et al. (2005) Infect. Immun. 73:935.
- Kuestner, R.E. et al. (2007) J. Immunol. 179:5462.
- Chang, S.H. and C. Dong (2007) Cell Res. 17:435.
- Afzali, B. et al. (2007) Clin. Exp. Immunol. 148:32.
- Fossiez, F. et al. (1996) J. Exp. Med. 183:2593.
- Yao, Z. et al. (1995) Immunity 3:811.
- Attur, M.G. et al. (1997) Arthritis Rheum. 40:1050.
- Ruddy, M.J. et al. (2004) J. Biol. Chem. 279:2559.
- Albanesi, C. et al. (1999) J. Immunol. 162:494.
- Witowski, J. et al. (2000) J. Immunol. 165:5814.
- Miyamoto, M. et al. (2003) J. Immunol. 170:4665.
- Ye, P. et al. (2001) Am. J. Respir. Cell Mol. Biol. 25:335.
- Laan, M. et al. (2001) Br. J. Pharmacol. 133:200.
- Starnes, T. et al. (2002) J. Immunol. 169:642.
- Jones, C.E. et al. (2002) Am. J. Respir. Cell Mol. Biol. 26:748.
- Yao, Z. et al. (1995) J. Immunol. 155:5483.
Long Name
Interleukin 17
Alternate Names
CTLA-8, CTLA8, IL-17A, IL17, IL17A
Entrez Gene IDs
Gene Symbol
IL17A
UniProt
Additional IL-17/IL-17A Products
Product Documents for Canine IL-17/IL-17A Antibody (665909)
Certificate of Analysis
To download a Certificate of Analysis, please enter a lot or batch number in the search box below.
Note: Certificate of Analysis not available for kit components.
Product Specific Notices for Canine IL-17/IL-17A Antibody (665909)
For research use only
Customer Reviews for Canine IL-17/IL-17A Antibody (665909)
There are currently no reviews for this product. Be the first to review Canine IL-17/IL-17A Antibody (665909) and earn rewards!
Have you used Canine IL-17/IL-17A Antibody (665909)?
Submit a review and receive an Amazon gift card!
$25/€18/£15/$25CAN/¥2500 Yen for a review with an image
$10/€7/£6/$10CAN/¥1110 Yen for a review without an image
Submit a review
Loading...
Associated Pathways