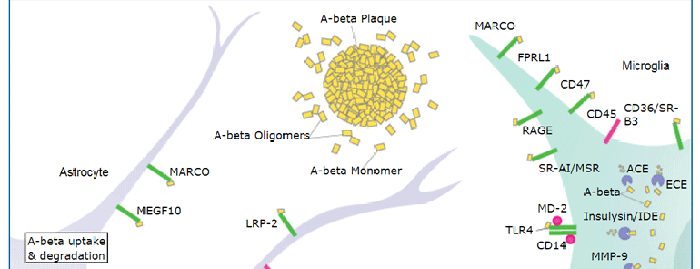
ApoE is a major protein component of serum LDL, VLDL, HDL, and chylomicrons. It is produced predominantly by hepatocytes, macrophages, and non-neuronal cells in the CNS. ApoE-containing particles transport triglycerides and cholesterol to peripheral tissues for cellular uptake and catabolism (1‑4). Mature human ApoE is a 37 kDa glycoprotein that consists of an N-terminal domain composed of four bundled alpha -helices, plus a hinge region and an extended alpha -helical C-terminal domain (2, 5). Its amphipathic nature and flexible structure enables it to adopt dramatically different conformations upon lipid association (2). ApoE is monomeric in lipid particles, although it forms oligomers when lipid-free (6). ApoE3 is the most abundant of the three common alleles in human; ApoE2 and ApoE4 differ by single aa substitutions (1). Mature human ApoE shares 71% aa sequence identity with mouse and rat ApoE. LDL receptor family proteins preferentially bind and internalize the lipid-bound form of ApoE with the exception of VLDLR which also efficiently internalizes lipid-free ApoE (7, 8). Lipoprotein uptake is facilitated by the initial binding of ApoE to cell surface heparan sulfate proteoglycans (HSPG) (9). Receptor/HSPG binding and lipid interactions primarily involve the N- and C-terminal regions of ApoE, respectively (2). Recycled lipid-free ApoE is formed into HDL particles through interactions with the lipid transporter ABCA1 (10). High cellular sterol content activates the nuclear hormone receptor LXR which promotes increased ApoE synthesis and increased sterol efflux, while low sterol content induces LDL R expression with increased sterol uptake and decreased ApoE production (11). ApoE3 dampens the TNF-alpha induced inflammatory response in vascular endothelial cells (12). In the CNS, ApoE blocks production of the amyloid A beta peptide by inhibiting the gamma -secretase cleavage of APP (13). It also complexes with A beta and promotes A beta internalization via LRP2 (14, 15).
Human Apolipoprotein E/ApoE Antibody (960318)
R&D Systems | Catalog # MAB41443

Key Product Details
Species Reactivity
Human
Applications
Immunohistochemistry
Label
Unconjugated
Antibody Source
Monoclonal Mouse IgG1 Clone # 960318
Loading...
Product Specifications
Immunogen
Human Apolipoprotein E/ApoE sythetic peptide
Accession # P02649
Accession # P02649
Specificity
Detects human Apolipoprotein E/ApoE in direct ELISAs.
Clonality
Monoclonal
Host
Mouse
Isotype
IgG1
Scientific Data Images for Human Apolipoprotein E/ApoE Antibody (960318)
Detection of Apolipoprotein E/ApoE in Human Brain.
Apolipoprotein E/ApoE was detected in immersion fixed paraffin-embedded sections of human brain (hippocampus) using Mouse Anti-Human Apolipoprotein E/ApoE Monoclonal Antibody (Catalog # MAB41443) at 5 µg/mL for 1 hour at room temperature followed by incubation with the Anti-Mouse IgG VisUCyte™ HRP Polymer Antibody (VC001). Tissue was stained using DAB (brown) and counterstained with hematoxylin (blue). Specific staining was localized to cytoplasm. View our protocol for IHC Staining with VisUCyte HRP Polymer Detection Reagents.Applications for Human Apolipoprotein E/ApoE Antibody (960318)
Application
Recommended Usage
Immunohistochemistry
5-25 µg/mL
Sample: Immersion fixed paraffin-embedded sections of human brain (hippocampus)
Sample: Immersion fixed paraffin-embedded sections of human brain (hippocampus)
Formulation, Preparation, and Storage
Purification
Protein A or G purified from hybridoma culture supernatant
Reconstitution
Reconstitute at 0.5 mg/mL in sterile PBS. For liquid material, refer to CoA for concentration.
Loading...
Formulation
Lyophilized from a 0.2 μm filtered solution in PBS with Trehalose. See Certificate of Analysis for details.
*Small pack size (-SP) is supplied either lyophilized or as a 0.2 µm filtered solution in PBS.
*Small pack size (-SP) is supplied either lyophilized or as a 0.2 µm filtered solution in PBS.
Shipping
Lyophilized product is shipped at ambient temperature. Liquid small pack size (-SP) is shipped with polar packs. Upon receipt, store immediately at the temperature recommended below.
Stability & Storage
Use a manual defrost freezer and avoid repeated freeze-thaw cycles.
- 12 months from date of receipt, -20 to -70 °C as supplied.
- 1 month, 2 to 8 °C under sterile conditions after reconstitution.
- 6 months, -20 to -70 °C under sterile conditions after reconstitution.
Calculators
Background: Apolipoprotein E/ApoE
References
- Martins, I.J. et al. (2006) Mol. Pschiatry 11:721.
- Hatters, D.M. et al. (2006) Trends Biochem. Sci. 31:445.
- Heeren, J. et al. (2006) Arterioscler. Thromb. Vasc. Biol. 26:442.
- Mahley, R.W. et al. (1984) J. Lipid. Res. 25:1277.
- Zannis, V.I. et al. (1984) J. Biol. Chem. 259:5495.
- Perugini, M.A. et al. (2000) J. Biol. Chem. 275:36758.
- Ruiz, J. et al. (2005) J. Lipid Res. 46:1721.
- Chroni, A. et al. (2005) Biochemistry 44:13132.
- Futamura, M. et al. (2005) J. Biol. Chem. 280:5414.
- Krimbou, L. et al. (2004) J. Lipid. Res. 45:839.
- Lucic, D. et al. (2007) J. Lipid Res. 48:366.
- Mullick, A.E. et al. (2007) Arterioscler. Thromb. Vasc. Biol. 27:339.
- Irizarry, M.C. et al. (2004) J. Neurochem. 90:1132.
- Naslund, J. et al. (1995) Neuron 15:219.
- Zerbinatti, C.V. et al. (2006) J. Biol. Chem. 281:36180.
Alternate Names
APOE
Gene Symbol
APOE
UniProt
Additional Apolipoprotein E/ApoE Products
Product Documents for Human Apolipoprotein E/ApoE Antibody (960318)
Certificate of Analysis
To download a Certificate of Analysis, please enter a lot or batch number in the search box below.
Note: Certificate of Analysis not available for kit components.
Product Specific Notices for Human Apolipoprotein E/ApoE Antibody (960318)
For research use only
Related Research Areas
Customer Reviews for Human Apolipoprotein E/ApoE Antibody (960318)
There are currently no reviews for this product. Be the first to review Human Apolipoprotein E/ApoE Antibody (960318) and earn rewards!
Have you used Human Apolipoprotein E/ApoE Antibody (960318)?
Submit a review and receive an Amazon gift card!
$25/€18/£15/$25CAN/¥2500 Yen for a review with an image
$10/€7/£6/$10CAN/¥1110 Yen for a review without an image
Submit a review
Protocols
Find general support by application which include: protocols, troubleshooting, illustrated assays, videos and webinars.
- Antigen Retrieval Protocol (PIER)
- Antigen Retrieval for Frozen Sections Protocol
- Appropriate Fixation of IHC/ICC Samples
- Cellular Response to Hypoxia Protocols
- Chromogenic IHC Staining of Formalin-Fixed Paraffin-Embedded (FFPE) Tissue Protocol
- Chromogenic Immunohistochemistry Staining of Frozen Tissue
- ClariTSA™ Fluorophore Kits
- Detection & Visualization of Antibody Binding
- Fluorescent IHC Staining of Frozen Tissue Protocol
- Graphic Protocol for Heat-induced Epitope Retrieval
- Graphic Protocol for the Preparation and Fluorescent IHC Staining of Frozen Tissue Sections
- Graphic Protocol for the Preparation and Fluorescent IHC Staining of Paraffin-embedded Tissue Sections
- Graphic Protocol for the Preparation of Gelatin-coated Slides for Histological Tissue Sections
- IHC Sample Preparation (Frozen sections vs Paraffin)
- Immunofluorescent IHC Staining of Formalin-Fixed Paraffin-Embedded (FFPE) Tissue Protocol
- Immunohistochemistry (IHC) and Immunocytochemistry (ICC) Protocols
- Immunohistochemistry Frozen Troubleshooting
- Immunohistochemistry Paraffin Troubleshooting
- Preparing Samples for IHC/ICC Experiments
- Preventing Non-Specific Staining (Non-Specific Binding)
- Primary Antibody Selection & Optimization
- Protocol for Heat-Induced Epitope Retrieval (HIER)
- Protocol for Making a 4% Formaldehyde Solution in PBS
- Protocol for VisUCyte™ HRP Polymer Detection Reagent
- Protocol for the Preparation & Fixation of Cells on Coverslips
- Protocol for the Preparation and Chromogenic IHC Staining of Frozen Tissue Sections
- Protocol for the Preparation and Chromogenic IHC Staining of Frozen Tissue Sections - Graphic
- Protocol for the Preparation and Chromogenic IHC Staining of Paraffin-embedded Tissue Sections
- Protocol for the Preparation and Chromogenic IHC Staining of Paraffin-embedded Tissue Sections - Graphic
- Protocol for the Preparation and Fluorescent IHC Staining of Frozen Tissue Sections
- Protocol for the Preparation and Fluorescent IHC Staining of Paraffin-embedded Tissue Sections
- Protocol for the Preparation of Gelatin-coated Slides for Histological Tissue Sections
- TUNEL and Active Caspase-3 Detection by IHC/ICC Protocol
- The Importance of IHC/ICC Controls
- Troubleshooting Guide: Immunohistochemistry
- View all Protocols, Troubleshooting, Illustrated assays and Webinars
Loading...
Associated Pathways