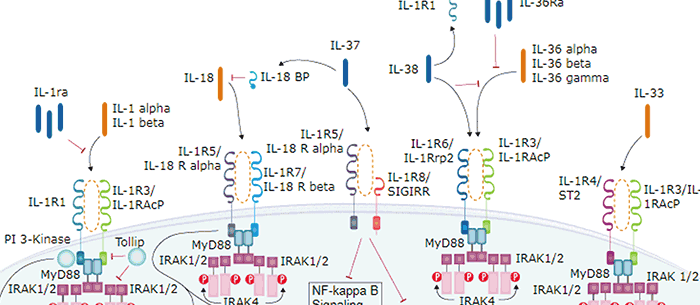
IL-33, also known as NF-HEV and DVS 27, is a 30 kDa proinflammatory protein that may also regulate gene transcription (1‑3). DVS 27 was identifed as a gene that is up‑regulated in vasospastic cerebral arteries (1). NF-HEV was described as a nuclear factor that is preferentially expressed in the endothelial cells of high endothelial venules relative to endothelial cells from other tissues (2). IL-33 was identified based on sequence and structural homology with IL-1 family cytokines (3). DVS 27, NF-HEV, and IL-33 share 100% amino acid sequence identity. IL-33 is constitutively expressed in smooth muscle and airway epithelia. It is up‑regulated in arterial smooth muscle, dermal fibroblasts, and keratinocytes following IL-1 alpha or IL-1 beta stimulation (1, 3). Similar to IL-1, IL-33 can be cleaved in vitro by caspase-1, generating an N-terminal fragment that is slightly shorter than the C-terminal fragment (3, 4). The N-terminal portion of full length IL-33 contains a predicted bipartite nuclear localization sequence and a homeodomain-like helix-turn-helix DNA binding domain. By immunofluorescence, full length IL-33 localizes to the nucleus in HUVECs and transfectants (2). The C-terminal fragment, corresponding to mature IL-33, binds and triggers signaling through mast cell IL-1 R4/ST2L, a longtime orphan receptor involved in the augmentation of Th2 cell responses (3, 5‑7). A ternary signaling complex is formed by the subsequent association of IL-33 and ST2L with IL-1R AcP (8). Stimulation of Th2 polarized lymphocytes with mature IL-33 in vitro induces IL-5 and IL-13 secretion (3). In vivo administration of mature IL-33 promotes increased production of IL-5, IL-13, IgE, and IgA, as well as splenomegaly and inflammatory infiltration of mucosal tissues (3). Full length and mature mouse IL-33 share approximately 55% and 90% aa sequence identity with human and rat IL-33, respectively. Mouse IL-33 shares less than 25% aa sequence identity with other IL-1 family proteins.
Mouse IL-33 Propeptide Antibody
R&D Systems | Catalog # AF5010

Key Product Details
Species Reactivity
Validated:
Mouse
Cited:
Mouse, Rat
Applications
Validated:
Western Blot, Immunocytochemistry
Cited:
Immunohistochemistry, Western Blot
Label
Unconjugated
Antibody Source
Polyclonal Sheep IgG
Loading...
Product Specifications
Immunogen
E. coli-derived recombinant mouse IL-33
Met1-Leu108
Accession # Q8BVZ5
Met1-Leu108
Accession # Q8BVZ5
Specificity
Detects mouse IL-33 Propeptide in direct ELISAs and Western blots. In direct ELISAs, less than 5% cross-reactivity with mature recombinant mouse IL-33 and recombinant human IL-33 Propeptide is observed.
Clonality
Polyclonal
Host
Sheep
Isotype
IgG
Scientific Data Images for Mouse IL-33 Propeptide Antibody
IL‑33 in bEnd.3 Mouse Cell Line.
IL‑33 was detected in immersion fixed bEnd.3 mouse endothelioma cell line using Sheep Anti-Mouse IL‑33 Propeptide Antigen Affinity-purified Polyclonal Antibody (Catalog # AF5010) at 10 µg/mL for 3 hours at room temperature. Cells were stained using the NorthernLights™ 557-conjugated Anti-Sheep IgG Secondary Antibody (red, upper panel; Catalog # NL010) and counterstained with DAPI (blue, lower panel). View our protocol for Fluorescent ICC Staining of Cells on Coverslips.
Detection of Mouse IL‑33 Propeptide by Western Blot.
Western blot shows lysates of EL-4 mouse lymphoblast cell line and mouse lactating mammary tissue. PVDF membrane was probed with 1 µg/mL of Sheep Anti-Mouse IL-33 Propeptide Antigen Affinity-purified Polyclonal Antibody (Catalog # AF5010) followed by HRP-conjugated Anti-Sheep IgG Secondary Antibody (Catalog # HAF016). A specific band was detected for IL-33 Propeptide at approximately 30 kDa (as indicated). This experiment was conducted under reducing conditions and using Immunoblot Buffer Group 8.Applications for Mouse IL-33 Propeptide Antibody
Application
Recommended Usage
Immunocytochemistry
5-15 µg/mL
Sample: Immersion fixed EL-4 mouse lymphoblast cell line and bEnd.3 mouse endothelioma cell line
Sample: Immersion fixed EL-4 mouse lymphoblast cell line and bEnd.3 mouse endothelioma cell line
Western Blot
1 µg/mL
Sample: EL‑4 mouse lymphoblast cell line and mouse lactating mammary tissue
Sample: EL‑4 mouse lymphoblast cell line and mouse lactating mammary tissue
Formulation, Preparation, and Storage
Purification
Antigen Affinity-purified
Reconstitution
Reconstitute at 0.2 mg/mL in sterile PBS. For liquid material, refer to CoA for concentration.
Loading...
Formulation
Lyophilized from a 0.2 μm filtered solution in PBS with Trehalose. *Small pack size (SP) is supplied either lyophilized or as a 0.2 µm filtered solution in PBS.
Shipping
Lyophilized product is shipped at ambient temperature. Liquid small pack size (-SP) is shipped with polar packs. Upon receipt, store immediately at the temperature recommended below.
Stability & Storage
Use a manual defrost freezer and avoid repeated freeze-thaw cycles.
- 12 months from date of receipt, -20 to -70 °C as supplied.
- 1 month, 2 to 8 °C under sterile conditions after reconstitution.
- 6 months, -20 to -70 °C under sterile conditions after reconstitution.
Calculators
Background: IL-33
References
- Onda, H. et al. (1999) J. Cereb. Blood Flow Metab. 19:1279.
- Baekkevold, E.S. et al. (2003) Am. J. Pathol. 163:69.
- Schmitz, J. et al. (2005) Immunity 23:479.
- Black, R.A. et al. (1989) J. Biol. Chem. 264:5323.
- Xu, D. et al. (1998) J. Exp. Med. 187:787.
- Lohning, M. et al. (1998) Proc. Natl. Acad. Sci. 95:6930.
- Dinarello, C.A. (2005) Immunity 23:461.
- Chackerian, A.A. et al. (2007) J. Immunol. 179:2551.
Long Name
Interleukin 33
Alternate Names
C9orf26, DVS27, IL33, NF-HEV
Gene Symbol
IL33
UniProt
Additional IL-33 Products
Product Documents for Mouse IL-33 Propeptide Antibody
Certificate of Analysis
To download a Certificate of Analysis, please enter a lot or batch number in the search box below.
Note: Certificate of Analysis not available for kit components.
Product Specific Notices for Mouse IL-33 Propeptide Antibody
For research use only
Related Research Areas
Citations for Mouse IL-33 Propeptide Antibody
 Powered by Bioz
Powered by Bioz
Customer Reviews for Mouse IL-33 Propeptide Antibody
There are currently no reviews for this product. Be the first to review Mouse IL-33 Propeptide Antibody and earn rewards!
Have you used Mouse IL-33 Propeptide Antibody?
Submit a review and receive an Amazon gift card!
$25/€18/£15/$25CAN/¥2500 Yen for a review with an image
$10/€7/£6/$10CAN/¥1110 Yen for a review without an image
Submit a review
Protocols
Find general support by application which include: protocols, troubleshooting, illustrated assays, videos and webinars.
- Appropriate Fixation of IHC/ICC Samples
- Cellular Response to Hypoxia Protocols
- ClariTSA™ Fluorophore Kits
- Detection & Visualization of Antibody Binding
- ICC Cell Smear Protocol for Suspension Cells
- ICC Immunocytochemistry Protocol Videos
- ICC for Adherent Cells
- Immunocytochemistry (ICC) Protocol
- Immunocytochemistry Troubleshooting
- Immunofluorescence of Organoids Embedded in Cultrex Basement Membrane Extract
- Immunohistochemistry (IHC) and Immunocytochemistry (ICC) Protocols
- Preparing Samples for IHC/ICC Experiments
- Preventing Non-Specific Staining (Non-Specific Binding)
- Primary Antibody Selection & Optimization
- Protocol for VisUCyte™ HRP Polymer Detection Reagent
- Protocol for the Fluorescent ICC Staining of Cell Smears - Graphic
- Protocol for the Fluorescent ICC Staining of Cultured Cells on Coverslips - Graphic
- Protocol for the Preparation and Fluorescent ICC Staining of Cells on Coverslips
- Protocol for the Preparation and Fluorescent ICC Staining of Non-adherent Cells
- Protocol for the Preparation and Fluorescent ICC Staining of Stem Cells on Coverslips
- Protocol for the Preparation of a Cell Smear for Non-adherent Cell ICC - Graphic
- R&D Systems Quality Control Western Blot Protocol
- TUNEL and Active Caspase-3 Detection by IHC/ICC Protocol
- The Importance of IHC/ICC Controls
- Troubleshooting Guide: Western Blot Figures
- Western Blot Conditions
- Western Blot Protocol
- Western Blot Protocol for Cell Lysates
- Western Blot Troubleshooting
- Western Blot Troubleshooting Guide
- View all Protocols, Troubleshooting, Illustrated assays and Webinars
Loading...
Associated Pathways