BDNF: Products
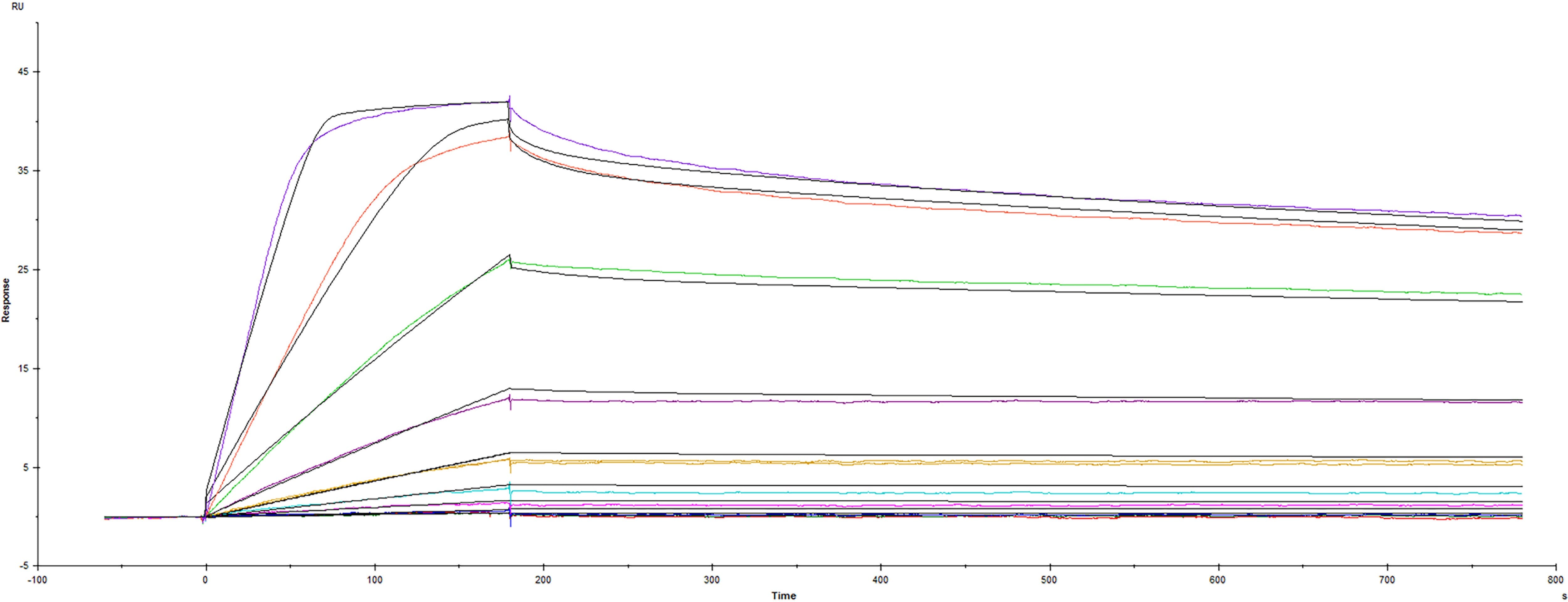
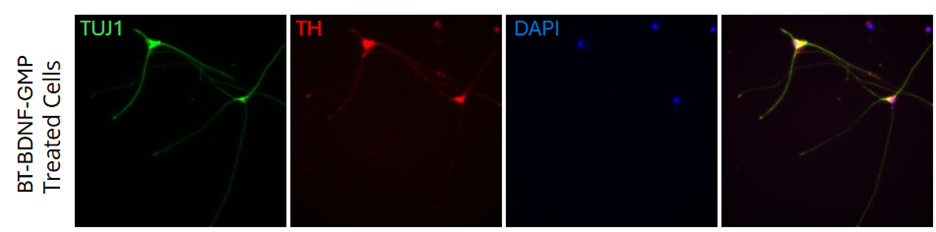
BDNF (Brain-Derived Neurotrophic Factor) is a secreted protein that regulates many aspects of neuronal development and function in the nervous system. It is produced as proBDNF in response to neuronal activity or inflammatory stimulation and is then cleaved before associating into a homodimer.
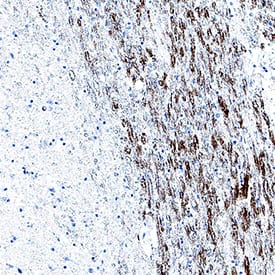
It is expressed throughout the nervous system as well as by fibroblasts, megakaryocytes/platelets, and smooth muscle cells. BDNF regulates neural stem cell survival and differentiation, axon/dendrite differentiation, synapse formation and maturation, and refinement of developing circuits. It is also a key regulator of synaptic plasticity and late-phase long-term potentiation. BDNF signals through the TrkB and NGF R/TNFRSF16 receptors. Secreted proBDNF can bind to NGF R/TNFRSF16 and induce long-term depression and cell apoptosis.
Products:
- Primary Antibodies
- Proteins & Enzymes
- ELISAs
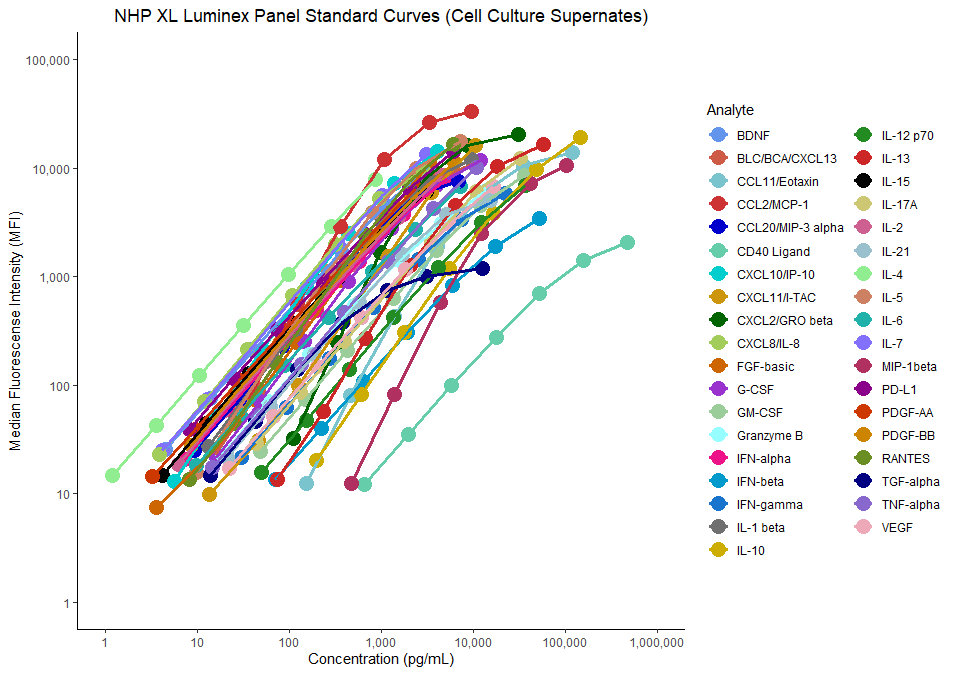
- Luminex Assays
- Simple Plex Assays
- cDNA Clones
- Peptides
- Proteome Profiler Antibody Arrays
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}